




Didžioji migracija Understand article
Išverstė Kauno regioninio inovacijų centro užsakymu, Kodėl ląstelės yra kaip gnu? Laura Spinney nagrinėja ląstelių migraciją ir organų formavimąsi, kaip pavyzdį naudodama mažytę ir permatomą zebrinę daniją.
Kiekvienais metais gegužės mėnesį didelės gnu bandos pajuda iš sausų Rytų Afrikos lygumų į miškus, ieškodamos maisto. Jos grįžta lapkričio mėnesį, kai lygumas palaisto trumpi lietūs, ir taip užbaigia kasmetinę migraciją. Visos gnu elgiasi pagal išlikimo instinktus, dėl kurių jos linksta prie kitų gnu. Susikūrus bandai, ji veikia kaip naujas vienetas, turintis savo taisykles. Ląstelės turi panašų bandos instinktą. Jos yra genetiškai užprogramuotos dirbti su kitomis ląstelėmis, tačiau nėra griežto taisyklių rinkinio, tiksliai nustatančio kiekvienos ląstelės elgesį grupėje. „Dabar mes suvokiame, kad daugelis veiksmų po apvaisinimo yra laisvi ir plastiški bei pagrįsti saviorganizacijos principais“, teigia Darren Gilmour iš Europos molekulinės biologijos laboratorijos Heidelberge, Vokietijoje.
Kaip vystymosi biologas Darren norėtų suprasti šiuos principus, nes masinis ląstelių judėjimas leidžia atkurti painias ir sudėtingas sistemas, pvz. kraujotakos arba kvėpavimo. Kadangi atrodo, kad migruojančių grupių ląstelės labai panašios į jų išreiškiamus genus, atskirų genų nuaktyvinimas arba veiklos susilpninimas blokuoja visos grupės migraciją. Tad tenka ieškoti subtilesnio būdo, kaip sugriauti ląstelių socialinius tinklus, stebint, kaip tai paveikia jų sugebėjimą migruoti ir sudaryti organus. Biologas teigia, kad tai iš esmės sociologija. „Mes norime žinoti, kas kam liepia ką ir kaip daryti.“

Paveikslėlio autorinės teisės priklauso Darren Gilmour, EMBL
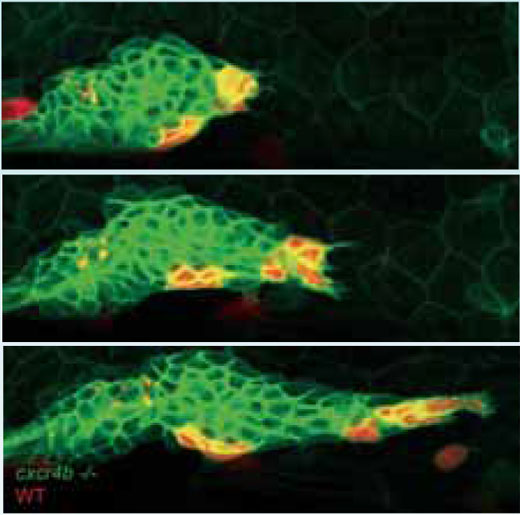
Darreno grupė tiria šiuos principus, nagrinėdama zebrinės danijos šoninės linijos gemalą – trumpalaikę sistemą, kurią galima matyti tik ankstyvo vystimosi metu, o jos funkcija yra aprūpinti embrioninę žuvį plaukų ląstelių organais – jutimo organais, leidžiančiais aptikti kitas žuvis, remiantis smulkiais vandens slėgio pasikeitimais. „Gemalas – tai daugiau nei 100 ląstelių grupė, šliaužianti po gyvūno oda kaip šliužas“, aiškina Darren. „Jis pradeda judėti iš už ausies link uodegos ir palieka maždaug dviejų ląstelių storio pėdsaką, pertraukiamą mažų ląstelių grupių, vėliau tapsiančių plaukų ląstelių organais”.
Pagal modelines sistemas, zebrinė danija yra atsakas į biologų maldas, nes ji permatoma ir todėl tinka ląstelių, pažymėtų fluorescentiniais proteinais, vaizdui gauti. Šoninės linijos sistemą yra gana lengva sukurti genetiškai. Be to, tai tikra dovana besidomintiems ląstelių migracija, nes ji naudoja signalizuojantį kelią, svarbų žmonių sveikatai. Cheminis signalas SDF1, atitinkantis žmonėms būdingus signalus, sukelia daug pasikeitimų gemalo ląstelėse. Tai padaroma prisijungiant prie receptoriaus, vadinamo Cxcr4b, kurį ląstelės perneša savo išorėje. Cxcr4b – tai proteinas, esantis skersai ląstelės membranos, o žmonių atveju, jis būtinas ŽIV infekcijai. Jis svarbus normaliam vystymuisi, bet kartu turi įtakos metastazei bei auglio ląstelių augimui.
Zebrinės danijos mutanto atveju, Darreno grupė nustatė, jog kai susilpnėja Cxcr4b signalas, gemalas netenka viso kryptingumo – nėra judesio į priekį ir šliaužiančio pėdsako. Be to, atrodo, kad statiška sistema nebegamina ląstelių grupių, tarsi jų gavimui reiktų pajudėti ir išstumti daugiau grupių. Tačiau vienas įdomiausių mutanto bruožų yra tas, kad net sustabdžius gemalo migraciją, ląstelės tebejuda, virsdamos viena per kitą gana atsitiktiniu būdu. ”Atrodo, tarytum ląstelės būtų sumišę”, teigia Darrenas. “Tarsi bezdžionės maiše”, o tai, anot mokslininko, yra raktas į visos sistemos veikimą
Norėdami išsiaiškinti, ar galima išgelbėti mutantą ir atstatyti judėjimą į priekį, tyrėjai sukūrė genetinę mozaiką, transplantuodami keletą normalių, „matančių“ ląstelių su išreikštu Cxcr4b receptoriumi ir galinčių atsiliepti į SDF1a, šiaip jau „aklą“ mutanto audinį. Šios normalios ir mutacinės ląstelės buvo pažymėtos atitinkamai raudonais ir žaliais fluorescentiniais proteinais, taigi Darren su komanda galėjo sekti ląstelių judėjimą. Labai greitai, per virstantį ląstelių judėjimą gemale, matančios ląstelės susirado kelią į priekį, ir gemalas vėl pajudėjo pirmyn.

Paveikslėlio autorinės teisės priklauso Petra Riedinger, EMBL
ląstelės (raudona) randa savo kelią
į priekį ir parodo migracijos kryptį
traukdamos mutantines ląsteles
(žalia) su savimi. Norėdami padidinti,
spauskite ant nuotraukos
Paveikslėlio autorinės teisės priklauso
Darren Gilmour, EMBL
Pasak Darreno, keista šiuose eksperimentuose buvo tai, kad ne visos jaučiančios, Cxcr4b išreiškiančios ląstelės rado kelią į priekį. Kai tik priekį pasiekė keletas ląstelių, atsitiktinis virstantis judesys sustojo, gemalas pajudėjo į priekį, o į priekį nepajudėjusios ląstelės liko, kur buvo „tarsi įšalę lede“. Jis mano, kad sistema savo stiprybę aptinka nestabilume. „Jis sukasi ir atlieka atsitiktinius judesius, taip užtikrindamas, kad galinčios vesti ląstelės, nuves ten, kur veda. Kai jos traukia, kitos sustingsta.”
Tačiau jeigu ne visos ląstelės tampa lyderėmis, kas apibrėžia lyderį? Kad ir kas tai būtų, tai tikrai nėra genų veiklos skirtumai. Darren teigia: „Ne genai pasako, kad tu esi lyderis, o tu sekėjas. Genai suteikia visoms ląstelėms receptorius, o tada jau viską lemia „tarpusavio kova“. Jis su bendradarbe Petra Haas mano, kad lyderiavimą labiausiai apibrėžia nestabili migruojančios grupės prigimtis. Atsitiktinis ląstelių vartymasis kai kurias laikinai padaro stipresnėmis nei kitos, nes jos yra arčiau signalo šaltinio. Tada jos prie jo prisikabina ir, jeigu sugeba išlaikyti savo receptoriaus aktyvumą, pajuda į priekį. Tačiau tai jokiais būdais nereiškia istorijos pabaigos.
Viena ląstelių migracijos teorija teigia, kad vadovaujančios ląstelės perneša sekėjus, pasyviai leidžiančius save paimti. Tačiau Darren šį modelį vertina skeptiškai, vadindamas jį keliautojų autostopu modeliu. Jis linkęs manyti, kad lyderiai nurodo kitiems tinkamą kryptį, o gemalo vidinė struktūra yra tokia, kad kiti gali aktyviai sekti. Jis mano, kad panašus principas gali būti pritaikytas visų organų vystymuisi. „Jeigu pažvelgsite į kraujagyslių išsišakojimą, manau, kad mažiausias ląstelių skaičius bus viršūnėje, jaučiančioje aplinkos signalus, o šių ląstelių tarpusavio veikla įtakoja kitų ląstelių elgesį. Kitos ląstelės neprivalo matyti pasaulio.“ Jis pateikia sakalo, tykančio varnėnų pulko, analogiją. Nebūtinai visi varnėnai turi matyti sakalą, kad tinkamai atsakytų pabėgdami – jiems tereikia pastebėti panišką kaimynų atsaką.
Istorija tampa dar sudėtingesne, nes paaiškėja, kad ne visi sekėjai elgiasi vienodai. Savo eksperimentuose Darren su komanda pastebėjo, kad gemalo priekiui judant pirmyn, ląstelės gale sulėtėja, taip versdamos audinį temptis. Doktorantas Guillaume Valentin nustatė ir kitą receptorių, kurį išreiškia šios galinės ląstelės, atsiliepiančios į SDF1a, tačiau įtakojančios kitokį elgesį nei iššauktas SDF1a/Cxcr4B – galinės ląstelės tampa šiek tiek nepriklausomos nuo Cxcr4b išreiškiančių lyderių ląstelių.
Gemalui judant į priekį, nukeliant ląstelių grupes į galą, bendras ląstelių judėjimas per sistemą yra į galą. Darren mano, kad judėdamos atgal ląstelės gali įjungti antrąjį receptorių. Jis teigia, kad galbūt dviejų ląstelių populiacijų su skirtingais receptoriais junginys gali sukelti vikšrinį judesį, kurio metu gemalo priekis ir galas judėtų į priekį pakaitomis. Tai įrodo kai kurie tyrimai su zebrinių danijų embrionais, kuriuose sumažėjusi antrojo receptoriaus veikla ir trūksta antrosios koordinuoto judesio dalies. „Vedantis kraštas nuolat traukia, tačiau negrįžta, todėl galiausiai pasiduoda ir parvirsta atgal.“
Jeigu jis teisus, naujieji rezultatai iškelia klausimą, kaip per dideles ląstelių grupes perduoti dinamišką informaciją. Būtina suprasti ne tik signalizavimą, bet ir jėgų sklidimą per ląstelių populiacijas, kalbant apie ląstelių tarpusavio sąveiką. Čia Darren ir tikisi pratęsti savo tyrimus. Tačiau šiuo metu jis džiaugiasi antrojo SDF1a receptoriaus atradimu zebrinės danijos gemale. Jis teigia: „Mintis, kad ląstelės gali jausti viduje, yra visiškai nauja. „Tai, ar toks pat principas taikytinas gnu, jau kitas klausimas.“
Resources
- Haas P, Gilmour D (2006) Chemokine signaling mediates self-organizing tissue migration in the zebrafish lateral line. Developmental Cell 10: 673-680. doi: 10.1016/j.devcel.2006.02.019
Institutions
Review
Man buvo malonu būti paprašytai peržiūrėti šį straipsnį, nes aš kitaip nebūčiau perskaičiusi jo taip kruopščiai. Tai labai gerai parašytas straipsnis, su daug spalvotų analogų, tokių kaip Afrikos kilmės gyvūnų migracijos susiejimas su besivystančių organizmų ląstelių migracija. Įspūdinga ne tik biologija su signaliniais baltymais, ŽIV infekcija, zebrine danija, vėžio ląstelėmis, mutuotomis “aklosiomis” ląstelėmis, bet taip pat įspūdingas yra mokslinių tyrimų grupės metodikos aprašymas, kuris skamba kaip detektyvinė istorija: pradedant teorijomis, atmetant modelius, pridedant naujų idėjų ir jas patikrinant.
Šis straipsnis labiausiai turėtų dominti biologijos mokytojus. Jis taip pat galėtų būti tikrai geras supratimo pasitikrinimas arba kaip papildoma medžiaga biologijos moksleiviams 16 metų ar daugiau. Kita vertus, jį galima naudoti ir diskusijose apie embriologiją, stuburinių vystymąsi, genų inžineriją, ryšius tarp ląstelių lyginant su ryšiais tarp organizmų, signalinius baltymus, vėžį, mutacijas, mokslinius tyrimus ar modeliavimo teorijos veiksmus.
Sue Howarth, Jungtinė Karalystė