




La grande migrazione Understand article
Traduzione di Anna Oddone. Perché le cellule si comportano come gli gnu? Laura Spinney indaga la migrazione delle cellule e la formazione degli organi attraverso il piccolo e trasparente pesce zebra.
Ogni anno in maggio vaste mandrie di gnu si spostano dalle aride pianure dell’Africa orientale verso le foreste, in cerca di cibo. Ritornano a novembre, quando le brevi piogge tornano a bagnare le pianure, completando cosi la loro migrazione annuale. Ogni gnu si comporta seguendo il suo istinto di sopravvivenza, che lo spinge a cercare la compagnia degli altri gnu. Una volta che il gregge si è formato, esso si comporta come una nuova unità, con regole proprie. Le cellule hanno lo stesso istinto di gruppo. Sono geneticamente programmate per operare con altre cellule, ma senza che ci sia un insieme definito di istruzioni che determini il comportamento specifico di ogni cellula all’interno del gruppo. “Ci rendiamo conto solo ora che molti degli eventi che hanno luogo dopo la fertilizzazione sono in una certa misura liberi, plastici e basati su principi di auto-organizzazione”, spiega Darren Gilmour dello European Molecular Biology Laboratory di Heidelberg, in Germania.
Come biologo dello sviluppo, Darren si propone di capire i principi che governano la migrazione di massa delle cellule, migrazione che dà origine a sistemi complessi quali la rete dei vasi sanguigni o il sistema respiratorio. Le cellule che compongono questi gruppi migratori sono molto simili tra loro in termini di espressione genetica: quindi sopprimere o ridurre l’attivita’ di singoli geni porta all’arresto della migrazione di tutto il gruppo. Darren deve quindi adottare un approccio diverso, trovando il modo di rompere la rete sociale cellulare e osservare in seguito come l’assenza di questa rete influisce sulla capacità delle cellule di migrare e formare organi. “Praticamente è sociologia,” spiega Darren. “Vogliamo capire chi dice a chi cosa fare, e come”.

Immagine cortesemente messa a disposizione da Darren Gilmour, EMBL
Il gruppo di Darren studia questi meccanismi osservando il primordio della linea laterale nel pesce zebra. Si tratta di una struttura transitoria, visibile solo all’inizio dello sviluppo, che fornisce all’embrione organi caratterizzati da cellule ciliate, ovvero organi sensoriali che permettono al pesce di percepire la presenza di altri pesci basandosi su cambiamenti minimi di pressione dell’acqua. “Il primordio è un insieme di più di 100 cellule che si spostano strisciando sotto la pelle dell’animale, come un lumacone”, spiega Darren. “Parte dietro l’orecchio e si dirige verso la coda. Migrando lascia dietro di sé una scia di circa due cellule di spessore, costellata qua e là da piccoli aggregati di cellule, che formeranno in seguito gli organi di cellule ciliate.”
Come organismo modello, il pesce zebra (Brachydanio rerio, anche noto come “zebrafish”, NdT) sembra essere una risposta alle preghiere dei biologi. Infatti il suo corpo trasparente permette una agevole osservazione degli organi interni e, tramite miscroscopia in vivo, permette di osservare cellule che siano state marcate con proteine fluorescenti. Il sistema della linea laterale è una vera benedizione per gli studiosi di migrazione cellulare perché, oltre ad essere relativamente facile da modificare tramite ingegneria genetica, sfrutta un “signaling pathway” (via di trasmissione dei segnali, NdT) importante per la salute umana e quindi largamente studiato. Un segnale chimico noto come SDF1, l’equivalente nel pesce zebra di un segnale chimico che si trova anche negli essere umani, genera una serie di cambiamenti nelle cellule che compongono il primordio. Produce questi cambiamenti quando si lega ad un recettore noto come Cxcr4b, che si trova sulla superficie esterna delle cellule. Cxcr4b è una proteina che risiede nella membrana delle cellule e, negli esseri umani, e’ necessaria perché si sviluppi l’infezione da HIV. Oltre a giocare un ruolo importante nello sviluppo, questa proteina e’ coinvolta nei processi di metastasi (ovvero il diffondersi delle cellule tumorali).
Il gruppo di Darren ha scoperto che in un mutante di pesce zebra che presenta una ridotta capacità di signaling mediato da Cxcr4b, il primordio perde la sua direzionalità: non avanza più e non lascia più dietro di sé una scia di cellule. Inoltre, questa struttura statica sembra perdere la capacità di produrre aggregati di cellule, quasi come se dovesse avanzare ed espellere tali aggregati al fine di poterne generare altri. Una delle caratteristiche più interessanti del mutante, tuttavia, sta nel fatto che, nonostante la migrazione del primordio sia bloccata, se si guarda attentamente si vede che le cellule continuano a muoversi individualmente, rotolano una sull’altra in maniera apparentemente disordinata. “Sembra quasi che le cellule siano confuse, come scimmie in un sacco”, dice Darren. E questa, aggiunge, è la chiave per capire come funziona il sistema.
Per verificare se potevano salvare il sistema e ripristinare il movimento in avanti del primordio, i ricercatori hanno creato un mosaico genetico traspiantando nel tessuto composto di cellule mutate (che sembrano essere “cieche”) alcune cellule non mutate (ovvero “normali”), che esprimono il ricettore Cxcr4b e sono quindi in grado di rispondere al segnale SDF1a. Le cellule normali e le cellule mutate sono state rispettivamente marcate con proteine fluorescenti rosse e verdi, per poter seguire il movimento delle cellule. Molto rapidamente, attraverso il movimento apparentemente disordinato delle cellule, le cellule “normali” si sono fatte strada verso la parte anteriore del gruppo e a quel punto il primordio ha ripreso ad avanzare.

Immagine cortesemente messa a disposizione da Petra Riedinger, EMBL
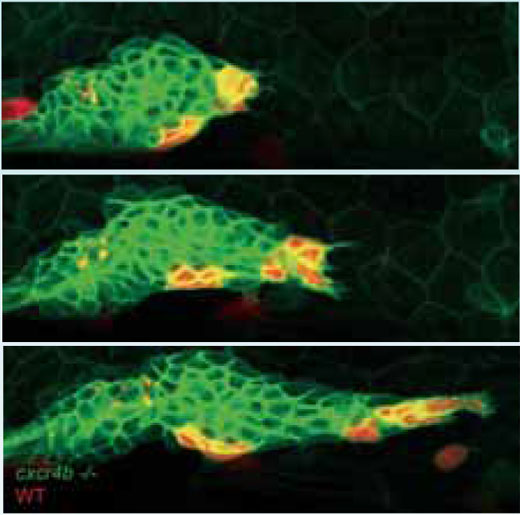
wild-type (in rosso) si fanno
strada verso l’estremità anteriore
del primordio e si estendono nella
direzione della migrazione,
trascinando con sé le cellule
mutanti (in verde). Cliccare sull’
immagine per ingrandire
Immagine cortesemente messa a
disposizione da Darren Gilmour,
EMBL
La cosa strana di questi esperimenti, racconta Darren, è che non tutte le cellule “normali” (quelle che esprimono Cxcr4b) sembrano farsi strada verso la parte anteriore del gruppo. Appena alcune di loro arrivano in testa al gruppo, il moto apparentemente disordinato delle cellule si ferma e il primordio riprende ad avanzare. Le cellule “normali” che non sono arrivate davanti in tempo rimangono dove sono, come se fossero congelate in quella posizione. “Pensiamo che il sistema trovi la sua forza nell’instabilità”, dice. “Il sistema gira intorno in maniera casuale e così facendo si assicura la possibilità che le cellule che possono dirigere il gruppo vengano a trovarsi in posizione per farlo. Non appena alcune di loro vi arrivano e cominciano a tirare, le altre si bloccano dove sono”.
Ma se non tutte le cellule “normali” finiscono per guidare il gruppo, che cosa distingue le cellule guida? Di qualsiasi cosa si tratti, non sembra risiedere in una diversa attività genetica. “Non sono i geni che dicono alle cellule, tu sei un leader, tu sei un gregario”, dice Darren. “I geni corredano tutte le cellule con il ricettore, dopodiché si tratta di una competizione tra cellule”. Darren e la sua collaboratrice Petra Haas pensano che la leadership sia definita dalla natura instabile del gruppo di cellule migratorie. Il ruotare disordinato delle cellule dà ad alcune di esse un vantaggio momentaneo rispetto ad altre, collocandole in prossimità della fonte del segnale. Una volta captato il segnale, se riescono a mantenere attivato il recettore, si spostano verso la parte anteriore del gruppo. Tuttavia la storia non finisce certamente qui.
Secondo una delle teorie della migrazione cellulare, le cellule-guida in qualche modo trascinano le gregarie, che passivamente si lasciano trainare. Tuttavia Darren ha qualche dubbio su quello che definisce il modello dell’autostoppista. Preferisce pensare che le cellule-guida indichino alle altre la direzione giusta e che il primordio sia organizzato internamente in maniera tale che le altre cellule seguano le cellule-guida attivamente. Crede inoltre che un principio simile valga per lo sviluppo di tutti gli organi. “Prendiamo ad esempio la ramificazione dei vasi sanguigni. Penso che ci sia all’estremità un numero limitato di cellule che percepiscono i segnali prodotti dall’ambiente circostante e che poi, tramite interazioni cellulari, influenzano in qualche modo il comportamento delle altre,” dice. “Le altre non hanno bisogno di vedere il mondo intorno.” Darren propone l’analogia con un falco a caccia di uno stormo di storni. Non occorre che tutti gli storni vedano il falco per reagire correttamente con la fuga, basta che percepiscano il panico e la risposta degli uccelli vicini.
Fin qui tutto bene, ma la storia si fa più complicata ancora, perché in realtà non tutte le cellule gregarie si comportano nello stesso modo. Negli esperimenti di microscopia in vivo, Darren e il suo gruppo hanno notato che, quando la parte anteriore del primordio si spinge in avanti, le cellule che stanno nella parte posteriore rallentano, causando così un allungamento del tessuto. Il dottorando Guillaume Valentin ha identificato un altro recettore, espresso dalle cellule posteriori, che risponde anch’esso al segnale chimico di SDF1a, ma che produce una risposta diversa da quella di Cxcr4b– fornendo così alle cellule posteriori una certa indipendenza da quelle anteriori, che invece esprimono Cxcr4b.
Quando il primordio avanza, depositando gruppi di cellule lungo il cammino, il movimento generale delle cellule all’interno del primordio è all’indietro. Darren ipotizza che, nel momento in cui si ritrovano nella parte posteriore, le cellule attivino questo secondo recettore. Forse, ipotizza, la concomitanza delle due popolazioni di cellule che esprimono due recettori diversi produce una specie di movimento del bruco, in cui la parte anteriore e la parte posteriore del primordio avanzano in modo alternato. Alcuni indizi che sembrano confermare questo tipo di processo vengono da esperimenti con embrioni di pesce zebra in cui l’attività del secondo recettore è ridotta: in questi embrioni la seconda fase di questo movimento coordinato è assente. “La parte anteriore tira e tira, ma la parte posteriore non segue, così alla fine la parte anteriore si arrende e ruzzola indietro,” dice.
Se Darren ha ragione, queste scoperte sollevano nuove domande su come l’informazione dinamica può essere trasmessa attraverso gruppi estesi di cellule. Diventa necessario capire non solo i processi di signalling, ma anche la propagazione delle forze attraverso popolazioni di cellule, in termini di interazioni cellulari. Questo è ciò che adesso Darren si ripropone di studiare. Per il momento, comunque, è elettrizzato dalla scoperta del secondo recettore per SDF1a nel primordio del pesce zebra. “L’idea che anche le cellule all’interno possano percepire il segnale è assolutamente nuova,” dice. Che poi lo stesso principio si possa applicare agli gnu, è tutta un’altra questione.
Resources
- Haas P, Gilmour D (2006) Chemokine signaling mediates self-organizing tissue migration in the zebrafish lateral line. Developmental Cell 10: 673-680. doi: 10.1016/j.devcel.2006.02.019
Institutions
Review
Sono felice che mi abbiano chiesto di fare una recensione di questo articolo, perché altrimenti non l’avrei forse letto con altrettanta attenzione. E’ scritto talmente bene, fa uso di numerose vivide analogie, come quella della migrazione delle cellule in un organismo in via di sviluppo con dei branchi di animali africani. Non solo la biologia trattata è affascinante, in quanto considera le proteine che agiscono come segnali chimici, l’infezione da HIV, il pesce zebra, le cellule tumorali e le cellule mutanti “cieche”, ma altrettanto affascinante è la descrizione della metodologia del gruppo di ricerca, che si legge come un romanzo giallo: le prime ipotesi, i modelli scartati, la formulazione di nuove idee e gli esperimenti per verificarle.
Questo articolo è di grande interesse per la maggior parte degli insegnanti di biologia. Può essere usato come esercizio di comprensione o fornire materiale aggiuntivo per studenti di biologia a partire dai 16 anni. Può anche essere usato per discussioni riguardanti l’embriologia, lo sviluppo dei vertebrati, l’ingegneria genetica, la comunicazione tra cellule rispetto alla comunicazione tra organismi, le proteine-segnale, il cancro, le mutazioni, le diverse fasi della ricerca scientifica o la formulazione di modelli e teorie.
Sue Howarth, Regno Unito