




La lumière synchrotron éclaire les origines obscures des orangs-outangs Understand article
Traduit par Paul Tafforeau. Paul Tafforeau du laboratoire de Géobiologie, Biochronologie et Paléontologie Humaine (université de Poitiers) et de I'Installation Européenne de Rayonnement Synchrotron (ESRF, Grenoble, France) nous explique ce que l’étude de dents fossiles par rayonnement X…

De nos jours, le groupe des hominoïdes (les grands singes sans queue) est représenté par seulement cinq genres: les gibbons, les orangs-outangs, les gorilles, les chimpanzés et notre propre espèce, Homo sapiens. Ce groupe a été très florissant pendant le Miocène entre 20 et 6 millions d’années comme le montre les nombreux genres fossiles (plus de 20) découverts en Afrique, Asie et Europe.
Les plus anciennes formes sont trouvées aux alentours de 20 MA en Afrique. Par la suite, on trouve les premiers hominoïdes hors d’Afrique à partir de 16.5 MA, puis de très nombreuses formes en Europe et en Asie entre 12 et 6 MA. Le groupe ne fera ensuite que décliner en diversité probablement pour des raisons de changements climatiques. Plusieurs hypothèses ont été proposées pour relier les différentes espèces fossiles et actuelles entre elles sur les plans phylogénétiques (les liens de parenté) et paléobiogéographiques (les migrations et centres de diversification).
L’hypothèse traditionnelle est celle d’un centre de diversification principal en Afrique pendant tout le Miocène, avec des migrations successives en Europe et en Asie associées à des évolutions locales sur ces trois continents (carte A).
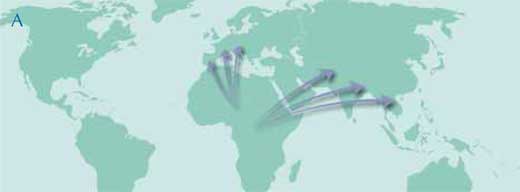
Image reproduite avec l’aimable autorisation de Petra Riedinger
Depuis quelques années, une nouvelle hypothèse est en train d’émerger. Elle propose également une origine du groupe en Afrique, puis une migration des formes primitives vers l’Asie couplée à une extinction progressive en Afrique, puis de nouvelles migrations depuis l’Asie vers L’Europe et l’Afrique (carte B). Selon cette seconde hypothèse, le centre principal de diversification des hominoïdes se trouverait en Asie et non pas en Afrique. Les fossiles clés pour pouvoir trancher entre ces deux scénarios devraient donc se trouver en Asie.
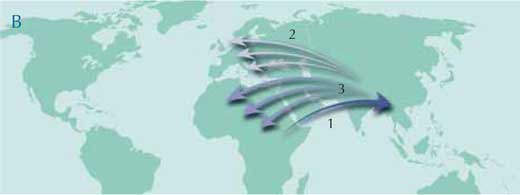
Image reproduite avec l’aimable autorisation de Petra Riedinger
En 2003, des recherches en Thaïlande ont permis de mettre à jour une nouvelle forme d’hominoïde âgée d’environ 12 MA (Chaimanee et al., 2003). Une vingtaine de dents isolée (figure 1A) attribuées à plusieurs individus mâles et femelles ont montré qu’il s’agit d’un hominoïde de grande taille avec un fort dimorphisme sexuel (Le mâle étant beaucoup plus gros que la femelle avec des canines plus développées) qui a été nommé cf. Lufengpithecus chiangmuanensis.
l’émail dentaire est le tissu le plus minéralisé qu’on puisse trouver chez les vertébrés, ce qui rend les dents très solides et très résistantes à l’érosion mécanique ou chimique. De plus, comme les dents contiennent très peu de matière organique pouvant être dégradée par les micro-organismes, elles sont généralement bien mieux préservées lors de la fossilisation que le reste de l’organisme, y compris les os. De ce fait, l’immense majorité du registre fossile des vertébrés est constituée de dents, ce qui est également vrai pour les hominoïdes. Heureusement pour les paléontologues, les dents présentent de nombreux caractères anatomiques liés à la fois à la phylogénie et à leurs fonctions, que ce soit pour l’alimentation ou en tant qu’attributs sexuels secondaires (i.e. les fortes canines chez les mâles chez de nombreux primates).
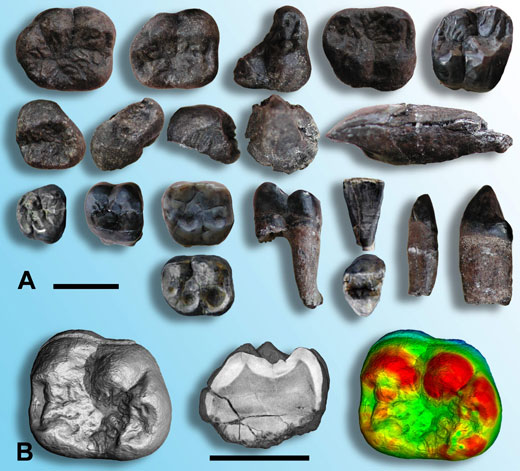
Image reproduite avec l’aimable autorisation de Paul Tafforeau
En plus d’une étude paléontologique classique portant sur la morphologie dentaire externe, la structure interne de dents de cf. Lufengpithecus chiangmuanensis a pu être étudiée de façon non destructive avec une qualité qui n’avait jamais été obtenue auparavant (Chaimanee et al., 2003; Tafforeau, 2004; Tafforeau et al., 2006). Grâce à l’utilisation de la ligne de lumière ID19 à l’Installation Européenne de Rayonnement Synchrotron (ESRF, Grenoble, France)w1 toutes les dents ont été imagées en trois dimensions par microtomographie à rayonnement X synchrotron. Ceci a permis de quantifier très précisément l’épaisseur et la distribution de l’émail (figure 1B) sans avoir à couper les dents. La combinaison de la paléontologie traditionnelle avec les analyses microtomographiques a permis de montrer que ce taxon fossile est plus proche des orangs-outangs actuels que tous les autres hominoïdes fossiles connus en Asie. Il pourrait donc prétendre au statut d’ancêtre des orangs-outangs.
En 2004, un nouveau fossile, âgé d’environ 7 MA a été découvert en Thaïlande. Il s’agit d’une très belle mandibule (figure 2A) qui a été nommée Khoratpithecus piriyai (Chaimanee et al., 2004). Sur la base de la taille générale, de la taille des canines et des structures osseuses, elle a été attribuée à un mâle. La comparaison de ce fossile avec les dents précédemment découvertes a montré que ces deux formes sont plus proches l’une de l’autre que de n’importe quel autre forme fossile ou actuelle, l’espèce la plus ancienne a donc été réattribuée à ce nouveau genre et renommée Khoratpithecus chiangmuanensis.
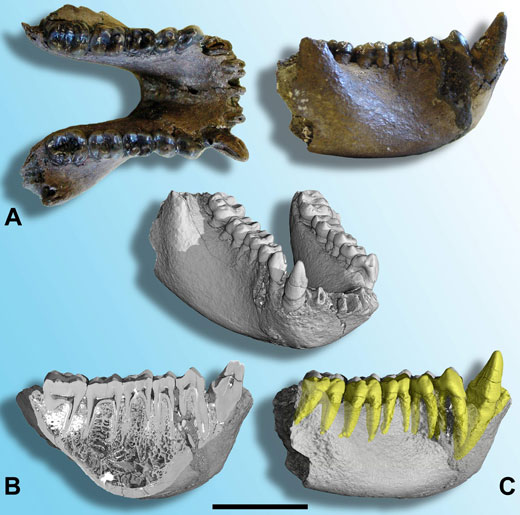
Image reproduite avec l’aimable autorisation de Paul Tafforeau
Cette mandibule de K.piriyai a également été imagée à l’ESRF par microtomographie à rayonnement X synchrotron, sur la ligne de lumière ID17 (ligne biomédicale). Cette expérience a permis de générer le premier scan de très haute qualité d’un fossile de grande taille d’hominoïde. Grâce à la préparation de ce jeu de données exceptionnelles il a été possible d’analyser la structure dentaire, d’étudier l’architecture osseuses de la mandibule (figure 2B) et d’extraire virtuellement toutes les dents du côté droit pour étudier la disposition et la taille des racines (figure 2C; Tafforeau, 2004; Tafforeau et al., 2006; Chaimanee et al., 2006).
Les données 3D des dents de K. chiangmuanensis ont été utilisées pour réaliser une reconstitution virtuelle des mâchoires de cet hominoïde (figure 3). Ce composite a permis de pousser plus loin les comparaisons entre les deux espèces de Khoratpithèques en prenant en compte la morphologie générale des mâchoires et les tailles relatives des dents entre elles. Cette comparaison a montré que les ressemblances entre ces deux taxons sont encore plus fortes que ce qu’on pensait, justifiant pleinement la réunion de ces deux espèces dans le même genre qui avait été faite sur la base d’une étude paléontologique classique.
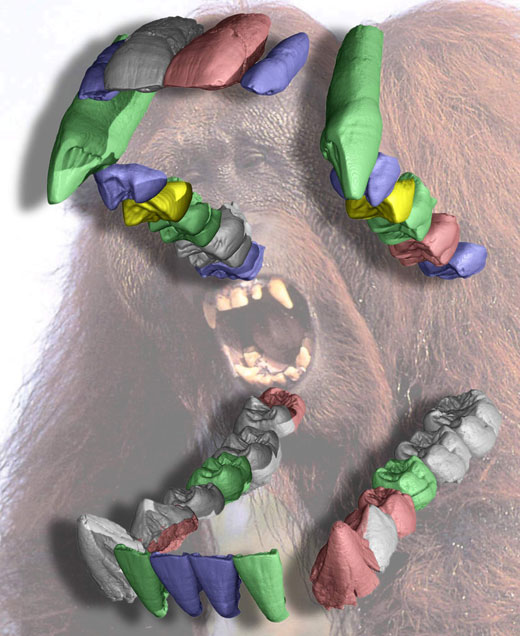
Image reproduite avec l’aimable autorisation de Paul Tafforeau
L’imagerie synchrotron appliquée à ces fossiles a révélé des caractères anatomiques impossibles à étudier sans détruire les fossiles. Ces analyses de pointe, couplées à une démarche paléontologique plus classique ont révélé que les Khorapithèques sont le groupe fossile connu le plus proche des orangs-outangs. Cependant, ces études ont également montré que K.piriyai (l’espèce la plus récente) bien que présentant beaucoup des caractères anatomiques typiques des orangs-outangs, présente également des caractères spécialisés qui lui sont spécifiques. Par exemple, K. piriyai a des canines en forme de dagues très différente de celles des orangs-outangs actuels et présente une troisième molaire extrêmement développée. En se basant sur ce qui est observé dans d’autres groupes, il est très peu probable que ce type de spécialisation puisse avoir évolué puis avoir subit une réversion pour revenir à un état plus primitif. K. piriyai, bien qu’étroitement apparenté aux orangs-outangs ne peut pas être considérée comme leur ancêtre direct.
K.chiangmuanensis est une espèce plus primitive avec moins de caractères spécialisés et correspond mieux à un ancêtre possible des orangs-outangs. Bien que de nouveaux fossiles soient nécessaires pour tester cette hypothèse, on peut donc proposer que la lignée des orangs-outangs s’enracine dans les formes anciennes du genre Khoratpithecus, celui-ci ayant continué à évoluer vers des formes plus spécialisées après la séparation des deux lignées.
Nous pouvons à présent revenir sur nos deux hypothèse de l’évolution des hominoïdes : le centre de diversification en Afrique ou en Asie. Les deux espèces de Khoratpithèques ainsi que de nombreux autres hominoïdes fossiles d’Asie présentent une bien plus forte diversité que les fossiles Africains connus pour la même période. De plus des espèces spécialisées et primitives ont été découvertes en Asie. La forte diversité et la vaste répartition géographique et temporelle des hominoïdes en Asie (de 16.5 millions d’années à nos jours) suggère fortement que l’Asie a été un centre important, voire même le centre principal, de diversification des hominoïdes au Miocène. Les recherches actuelles apportent donc de plus en plus de poids à cette nouvelle hypothèse de l’évolution des hominoïdes : nos ancêtres hominoïdes seraient apparus en Afrique, se seraient ensuite diversifiés principalement en Asie, puis auraient repeuplé l’Afrique et l’Europe.
References
- Chaimanee Y et al. (2003) A Middle Miocene hominoid from Thailand and orangutan origins. Nature 422: 61-65. doi:10.1038/nature01449.
- Chaimanee Y et al. (2004) A new orang-utan relative from the Late Miocene of Thailand. Nature 427: 439-441. doi:10.1038/nature02245.
- Chaimanee Y et al. (2006) Khoratpithecus piriyai, a late Miocene hominoid of Thailand. American Journal of Physical Anthropology 131: 311-323. doi:10.1002/ajpa.20437
- Tafforeau P (2004) Aspects phylogénétiques et fonctionnels de la microstructure de l’émail dentaire et de la structure tridimensionnelle des molaires chez les primates fossiles et actuels : apports de la microtomographie à rayonnement X synchrotron. Ph.D. Thesis, Université de Montpellier II, France
- Tafforeau P et al. (2006) Applications of X-ray synchrotron microtomography for non-destructive 3D studies of paleontological specimens. Applied Physics A, Materials Science & Processing 83: 195-202. doi:10.1007/s00339-006-3507-2
Web References
- w1 – L’installation Européenne de Rayonnement Synchrotron (ESRF) est un institut international qui fait fonctionner et développe la plus puissante source de rayonnement synchrotron en Europe, avec plus de 18 pays membres. Plus de 5000 chercheurs viennent à l’ESRF tous les ans pour utiliser la lumière synchrotron et les instruments qui y sont associés.
- l’ESRF est un membre de l´EIROforum, une collaboration de sept organisations de recherches intergouvernementales européennes, qui est également l’éditeur de Science in School.
Resources
- Des exemples d’applications de l’usage de la lumière synchrotron à l’ESRF sont disponibles (here).
- Tafforeau P et al. (2005) Synchrotron Radiation Microtomography: A Tool for Paleontology. ESRF Newsletter 42: 22-23.
- Pour des informations sur l’université de Poitiers: www.univ-poitiers.fr
Institutions
Review
J’ai toujours été fasciné par les connaissances sur l’évolution de l’Homme. Les questions comme d’où venons nous ? et Qui a été notre ancêtre commun ? sont souvent posées à la fois par les élèves et les scientifiques. Cet article décrit une méthode qui aide à renforcer une des hypothèses de l’évolution des hominoïdes : nos ancêtres seraient originaires d’Afrique, ils se seraient ensuite diversifié en Asie, puis auraient finalement repeuplé l’Afrique et l’Europe. Cet article peut être utilisé pour des cours de sciences et vie de la Terre au plus haut niveau, pour des élèves âgés de 16 à 18 ans. Il pourrait être très utile lors de l’enseignement de l’évolution en général, et de l’évolution humaine en particulier. Cet article pourrait également être utilisé dans les cours de physique afin de montrer comment l’utilisation poussée des rayons X peut être un outil très précieux pour les biologistes.
Sølve Tegnér Stenmark, Norvège