




Promieniowanie synchrotronowe rzuca nowe światło na niejasne pochodzenie orangutanów Understand article
Tłumaczenie Izabela Szczupakowska. Paul Tafforeau z Uniwersytetu Poitiers i Europejskiego Centrum Promieniowania Synchrotronowego (ESRF) z Grenoble (Francja, tłumaczy jak badania skamieniałości zębów za pomocą synchrotronowego promieniowania X mogą wyjaśnić ewolucję orangutanów – i…

Obecnie Małpy Człekokształtne są reprezentowane przez pięć rodzajów: gibbony (rodzina gibbonowate) i orangutany, goryle, szympansy oraz nasz własny gatunek, Homo sapiens (rodzina człowiekowate). W epoce miocenu, 20-6 milionów lat temu, grupa ta była dużo bardziej zróżnicowana, poparte jest to analizą wielu (około 20) rodzajów skamieniałości odkrytych na terenie Afryki, Azji i Europy.
Najstarsze Małpy Człekokształtne pojawiły się około 20 milionów lat temu w Afryce. Wiadomo, że poza Afrykę pierwsze Małpy Człekokształtne wywędrowały około 16,5 miliona lat temu, i że wiele ich różnych form żyło w Europie i Azji 12 – 6 miliona lat temu. Następnie zmniejszyła się różnorodność tej grupy, prawdopodobnie w skutek zmian klimatycznych. Zostało zaproponowanych wiele hipotez wyjaśniających filogenetyczne (ewolucyjne) związki między skamieniałościami, a istniejącymi obecnie gatunkami oraz opisujących ich paleogeografię – to znaczy ich migrację geograficzną i różnicowanie.
Obowiązująca hipoteza mówi, że podczas epoki Miocenu w Afryce zachodziło różnicowanie, po którym nastąpiła migracja do Europy i Azji i dalsza lokalna ewolucja na trzech kontynentach (mapa A).
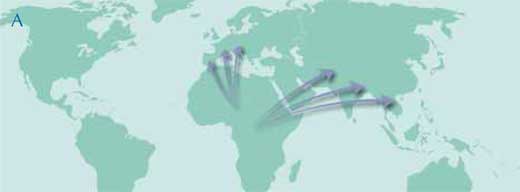
Ilustracja dzięki uprzejmości Petra Riedinger
Jednakże w ostatnich latach pojawiła się nowa hipoteza. Ona równiez proponuje Afrykę jako kontynent pochodzenia Małp Człekokształtnych, ale według niej prymitywne formy emigrowały w stronę Azji. Po stopniowym wyginięciu w Afryce i Europie populacje zostały odbudowane poprzez osobniki migrujące z Azji (mapa B). Według tej hipotezy to właśnie Azja, a nie Afryka była głównym miejscem różnicowania Małp Człekokształtnych. Kluczowe skamieniałości dla potwierdzenia tych dwóch scenariuszy powinny zatem być znalezione na tym właśnie kontynencie.
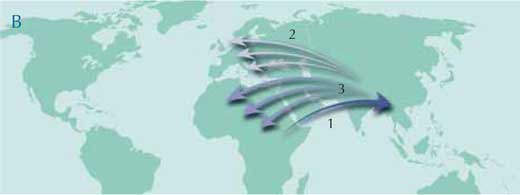
Ilustracja dzięki uprzejmości Petra Riedinger
W roku 2003 badania w Tajlandii doprowadziły do odkrycia skamieniałości nieznanego gatunku Małpy Człekokształtnej z przed około 12 milionów lat (Chaimanee et al., 2003). Około 20 wyizolowanych zębów (Ilustracja 1A), przypisanych do kilkunastu osobników, samców i samic, potwierdziło że odkryty gatunek był dużą Małpą Człekokształtną z silnie zaznaczonym dymorfizmem płciowym – samiec był dużo większy od samicy i miał bardziej rozwinięte kły. Gatunek ten został nazwany Lufengpithecus chiangmuanensis.
Szkliwo jest najbardziej zmineralizowaną tkanką kręgowców, w rezultacie zęby są bardzo mocne i odporne na urazy mechaniczne i czynniki chemiczne. Zęby zawierają również bardzo niewiele materii organicznej, która mogłaby być rozłożona przez mikroorganizmy. Te fakty oznaczają, że zęby są często lepiej zachowane podczas procesu fosylizacji niż reszta organizmu, w tym kości. Faktycznie ogromną większość skamieniałości kręgowców stanowią zęby i większość badań paleontologicznych nad szczątkami kręgowców (w tym Małp Człekokształtnych) jest oparta na ich analizie. Na szczęście dla paleontologów, zęby mają wiele cech anatomicznych, które pozwalają wnioskować o ich ewolucji i pełnionej funkcji. Określają zarówno rodzaj spożywanego pokarmu, jak i na ich podstawie można określić płeć osobnika (na przykład wiele samców z rzędu Naczelnych posiada wyjątkowo dobrze rozwinięte kły).
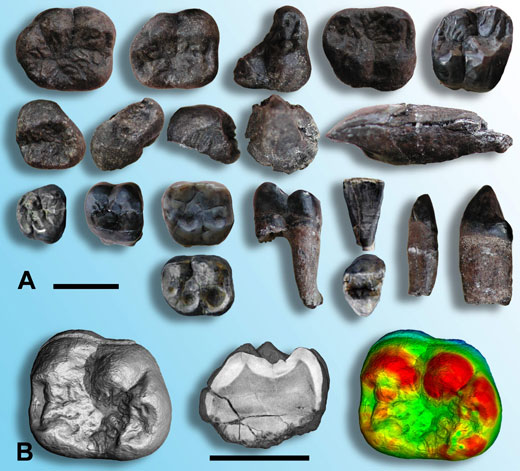
Ilustracja dzięki uprzejmości Paul Tafforeau
Oprócz przeprowadzenia tradycyjnych badań paleontologicznych zewnętrznej struktury zębów L. chiangmuanensis, zbadano również ich budowę wewnętrzną w sposób niedestrukcyjny, dostarczając danych o niedościgłej jakości (Chaimanee et al., 2003; Tafforeau, 2004; Tafforeau et al., 2006). W Europejskim Centrum Promieniowania Synchrotronowego (ESRF)w1 w Grenoble (Francja) zęby zostały zobrazowane w trzech wymiarach przy użyciu synchrotronowej mikrotomografii promieniowania X. Technika ta umożliwiła ilościowe i precyzyjne określenie grubości i dystrybucji szkliwa (Ilustracja 1B) bez potrzeby cięcia zęba. Wyniki badań tradycyjnej paleontologii oraz analiz mikrotomograficznych udowodniły, że skamieliny nieznanego gatunku są bardziej podobne do współczesnych orangutanów, niż do jakichkolwiek skamielin Małp Człekokształtnych z Azji. Zostało więc zaproponowane, że L. chiangmuanensis mógł być przodkiem orangutanów.
W 2004 roku, młodsze skamieliny, z przed około 7 milionów lat, zostały odkryte w Tajlandii. Idealnie zachowana żuchwa (szczęka dolna; Ilustracja 2A) została przypisana do nowego gatunku, nazwanego: Khoratpithecus piriyai (Chaimanee et al., 2004). Na podstawie ogólnej wielkości, wielkości kłów oraz morfologii kości, szczęka ta została uznana za pochodzącą od samca. Późniejsze porównania tej skamieliny z zębami L. chiangmuanensis potwierdziły, że obie te formy były do siebie bardziej podobne i bliżej ze sobą spokrewnione, niż z jakąkolwiek inną skamieliną bądź istniejącym gatunkiem. Starszy gatunek L. chiangmuanensis, został zatem przypisany do nowego rodzaju i przemianowany na Khoratpithecus chiangmuanensis.
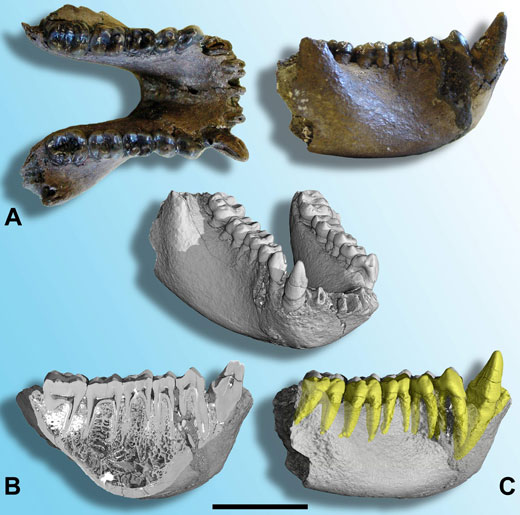
Ilustracja dzięki uprzejmości Paul Tafforeau
Szczęka dolna K.piriyai została również zobrazowana w ESFR przy użyciu synchrotronowej mikrotomografii promieniowania X, dostarczając pierwszego, doskonałej jakości obrazu tak dużej skamieliny Małpy Człekokształtnej. Posługując się tymi wyjątkowymi danymi, możliwe było analizowanie struktury zębów i budowy kości (Ilustracja 2B) oraz stworzenie wizualizacji zębów z prawej strony żuchwy w celu ocenienia wielkości i kształtu korzeni (Ilustracja 2C; Tafforeau, 2004; Tafforeau et al., 2006; Chaimanee et al., 2006).
Trójwymiarowy obraz zębów K. chiangmuanensis został użyty do właściwej rekonstrukcji żuchwy (Ilustracja 3), która pozwoliła na dalsze porównywanie ogólnej morfologii szczęk i relatywnego rozmiaru zębów dwóch gatunków Khoratpithecus. Badania te potwierdziły, że podobieństwa między tymi dwoma gatunkami były nawet większe niż początkowo przypuszczano i całkowicie uzasadniają ich klasyfikację w obrębie jednego rodzaju, co było dokonane na podstawie badań paleontologicznych.
Ilustracja dzięki uprzejmości Paul Tafforeau
Dzięki użyciu synchrotronowego promieniowania X w obrazowaniu skamielin stało się możliwym odkrycie anatomicznej budowy, bez jednoczesnego niszczenia skamieliny. Te przełomowe analizy, w połączeniu z tradycyjnymi badaniami paleontologicznymi pokazały, że Khorapithecus jest najbliżej spokrewnionym rodzajem z orangutanami. Niemniej, badania wykazały, iż K. piriyai (ten nowszy gatunek) posiadał wiele anatomicznych cech typowych dla orangutanów i jednocześnie wykazywał cechy różnicujące. Między innymi, K. piriyai posiadał niektóre zęby bardzo odmienne od tych jakie posiadają współczesne orangutany, sztyletowate kły oraz wyjątkowo duże trzecie zęby trzonowe. Z tego co wiadomo o rodowodzie tych małp jest nieprawdopodobne, że tak wiele cech najpierw wyewoluowało by następnie zaniknąć.
Dlatego K. piriyai chociaż spokrewniony ze współczesnymi orangutanami, to prawie na pewno nie jest ich bezpośrednim przodkiem. Natomiast K. chiangmuanensis, ten starszy gatunek, z mniej wyspecjalizowanymi zębami jest bardziej prawdopodobnym przodkiem orangutanów. Pomimo, że nowe skamieliny są potrzebne, aby potwierdzić tę hipotezę, prawdopodobnie linia orangutanów pochodzi od prymitywnych form rodzaju Khoratpithecus i że dwie gałęzie ewoluowały w różny sposób, dając początek zarówno orangutanom jak i późniejszemu rodzajowi Khoratpithecus, np. gatunkowi K. piriyai.
Możemy teraz powrócić do naszych dwóch hipotez ewolucji Małp Człekokształtnych: afrykańskiej i azjatyckiej. Dwa gatunki Khoratpithecus, jak również wiele innych skamielin azjatyckich Małp Człekokształtnych, wykazuje wyższe zróżnicowanie niż afrykańskie Małpy Człekokształtne w tym samym okresie. Co więcej, zarówno bardzo wyspecjalizowane jak i prymitywne gatunki zostały znalezione w Azji. Tak więc, wysoki poziom zróżnicowania oraz szeroki zasięg geograficzny w geologicznych warstwach, azjatyckich Małp Człekokształtnych (od 16,5 miliona lat temu do teraz) wyraźnie sugeruje, że Azja była ważnym, a być może najważniejszym, centrum różnicowania mioceńskich Małp Człekokształtnych. Zatem, coraz więcej nowoczesnych badań paleontologicznych potwierdza najnowszą hipotezę ewolucji Małp Człekokształtnych: nasz wspólny przodek pochodził z Afryki, potem różnicował się w Azji i odbudował populację w Afryce i Europie.
References
- Chaimanee Y et al. (2003) A Middle Miocene hominoid from Thailand and orangutan origins. Nature 422: 61-65. doi:10.1038/nature01449
- Poniżej możesz pobrać artykuł bezpłatnie. Możesz się także już dziś zapisać na stronie Nature: www.nature.com/subscribe
- Chaimanee Y et al. (2004) A new orang-utan relative from the Late Miocene of Thailand. Nature 427: 439-441. doi:10.1038/nature02245
- Poniżej możesz pobrać artykuł bezpłatnie. Możesz się także już dziś zapisać na stronie Nature: www.nature.com/subscribe
- Chaimanee Y et al. (2006) Khoratpithecus piriyai, a late Miocene hominoid of Thailand. American Journal of Physical Anthropology 131: 311-323. doi:10.1002/ajpa.20437
- Tafforeau P (2004) Aspects phylogénétiques et fonctionnels de la microstructure de l’émail dentaire et de la structure tridimensionnelle des molaires chez les primates fossiles et actuels : apports de la microtomographie à rayonnement X synchrotron. Ph.D. Thesis, Université de Montpellier II, France
- Tafforeau P et al. (2006) Applications of X-ray synchrotron microtomography for non-destructive 3D studies of paleontological specimens. Applied Physics A, Materials Science & Processing 83: 195-202. doi: 10.1007/s00339-006-3507-2
Web References
- w1 – Europejskie Centrum Promieniowania Synchrotronowego (ESRF) jest międzynarodowym centrum, które obsługuje i rozbudowuje najsilniejsze źródło światła synchrotronowego w Europie we współpracy z 18 krajami europejskimi. Ponad 5000 naukowców corocznie korzysta w ESRF ze źródła światła i związanych z nim urządzeń.
- ESRP jest członkiem EIROforum, siedmiu współpracujących, pozarządowych, europejskich organizacji naukowych oraz wydawców Science in School.
Resources
- Wyjaśnienie pojęcia światła synchrotronowego dostępne jest tutaj.
- Artykuł: Tafforeau P et al. (2005) Synchrotron Radiation Microtomography: A Tool for Paleontology. ESRF Newsletter 42: 22-23.
- Informacje o Uniwersytecie Poitiers: www.univ-poitiers.fr
Institutions
Review
Od zawsze byłem zafascynowany nauką o ewolucji gatunku ludzkiego. Pytania typu: „Skąd pochodzimy?”, „Kto był naszym wspólnym przodkiem?” są często zadawane przez uczniów i naukowców. Ten artykuł opisuje metodę, która wstępnie potwierdziła hipotezę ewolucji Małp Człekokształtnych, według której nasz wspólny przodek pochodził z Afryki, a następnie różnicował się w Azji i odbudował populację w Afryce i Europie. Ten artykuł może być użyty na lekcji biologii o poziomie rozszerzonym, dla uczniów w wieku 16-18. Byłby również bardzo przydatny podczas nauczania ewolucji, w szczególności ewolucji gatunku ludzkiego. Może być również użyty na lekcjach fizyki, aby pokazać w jaki sposób nowoczesne użycie promieniowania X może być wartościowym narzędziem dla biologa.
Sølve Tegnér Stenmark, Norwegia